Сцепленное с полом наследование птиц. Наследование признаков, сцепленных с полом. Вопросы и задания для повторения
Задача 1
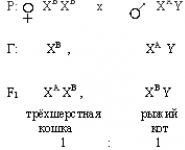
Известно, что «трехшерстные» кошки - всегда самки. Это обусловлено тем, что гены черного и рыжего цвета шерсти аллельны и находятся в Х – хромосоме. Ни один из них не доминирует, а при сочетании рыжего и черного цвета формируются «трехшерстные» особи.
1. Какова вероятность получения в потомстве «трехшерстных» котят от скрещивания «трехшерстной» кошки с черным котом?
2. Какое потомство можно ожидать от скрещивания черного кота с рыжей кошкой?
Решение:
Интересное сочетание: гены черного и рыжего цвета не доминируют друг над другом, а в сочетании дают «трехшерстную» масть. Здесь наблюдается кодоминирование (взаимодействие генов). Возьмем: А – ген отвечающий за черный цвет, В – ген отвечающий за рыжий цвет; гены А и В равнозначны и аллельны (А=В), но эти гены находятся в Х – хромосоме. Поэтому мы обозначаем ген черного цвета Х A , ген рыжего цвета – Х В.
По условия пункта 1 скрещиваются трехшерстная кошка с черным котом.
(вероятные фенотипы)
По условиям пункта 2 скрещиваются рыжая кошка с черным котом:

(вероятные фенотипы)
При решении задачи использовали закон чистоты гамет и сцепленное с полом наследование. Взаимодействие генов: кодоминирование. Вид скрещивания: моногибридное.
У человека гемофилия передается как рецессивный сцепленный с Х – хромосомой признак
Задача 2
Классическая гемофилия передается как рецессивный сцепленный с Х – хромосомой признак.
1. Мужчина, больной гемофилией, жениться на женщине, не имеющей этого заболевания. У них рождаются нормальные дочери и сыновья, которые все вступают в брак с не страдающими гемофилией лицами. Обнаружится ли у внуков вновь гемофилия, и какова вероятность появления больных в семье дочери или сына?
2. Мужчина, больной гемофилией, вступает в брак с нормальной женщиной, отец которой страдал гемофилией.
Определите вероятность рождения в этой семье здоровых детей.
Решение:

По условиям пункта 1 ясен генотип больного мужчины: Х h . Так как женщина не страдает гемофилией, у нее обязательно должен быть доминантный ген «нормы» - Х H . Второй ген женщины также доминантный (Х H), в генотип женщины Х H Х H . Генотипы детей от такого брака:

Иначе говоря, все мальчики будут здоровы, гена гемофилии у них не будет, а все девочки будут гетерозиготными – в рецессиве у них будет ген гемофилии.
Если все мальчики в последствии вступят в брак со здоровыми в отношении гемофилии лицами (Х H Х H), гемофилия у внуков не проявится. Если дочери (Х H Х h) вступят в брак со здоровыми мужчинами (Х H Y), вероятность проявления гемофилии у внуков будет равна1/4, или 25%. По полу это будут мальчики:

По условиям пункта 2 в брак вступает больной мужчина (генотип X h Y) с женщиной, не страдающей болезнью. Следовательно, у женщины один ген обязательно «норма» – Х H . Но второй ген из этой пары у нее должен быть геном гемофилии – Х h , так как отец этой женщины страдал гемофилией, а женщина получает всегда одну Х – хромосому от матери, а вторую – от отца. Генотип женщины – Х H Х h . Вероятность рождения здоровых детей в этой семье равна 1/2. Девочки с генотипом Х h Х h погибают.

Задача 3
У человека классическая гемофилия наследуется как сцепленный с Х – хромосомой рецессивный признак. Альбинизм (отсутствие пигментации) обусловлен аутосомным рецессивным геном. У одной супружеской пары, родился сын с обеими аномалиями. Какова вероятность того, что у второго сына в этой семье проявится также обе аномалии одновременно?
Решение:

По условиям задачи оба родителя нормальны, следовательно, у них обязательно есть по доминантному гену из каждой пары Х H и А. Сын имеет обе аномалии, его генотип Х h Yаа. Х – хромосому с геном гемофилии он мог унаследовать только от матери. Один из генов альбинизма сын получил от матери, другой - от отца. Таким образом, генотип матери Х H Х h Аа, генотип отца Х H YАа. При таком браке вероятны генотипы детей:

Вероятность того, что следующий ребенок будет сыном, равна 1/2. Из числа сыновей лишь 1/8 может иметь одновременно обе аномалии. Для вычисления окончательного результата вероятности перемножаются: 1/2 х 1/8=1/16.
Гипертрихоз передается как сцепленный с Y - хромосомой признак
Задача 4
Гипертрихоз (вырастание волос на краю ушной раковины) передается через Y – хромосому, а полидактилия (шестипалость) – как доминантный аутосомный ген. В семье, где отец имел гипертрихоз, а мать – полидактилию, родилась нормальная в отношении обоих признаков дочь. Какова вероятность того, что следующий ребенок в этой семье будет также без обеих аномалий?
Решение:
В условиях задачи ген гипертрихоза обозначим звездочкой (*), находящейся в Y*– хромосоме, в Х – хромосоме нет гена аллельного гену гипертрихозу:

Так как отец имел гипертрихоз и был пятипалым, его генотип XY*aa. У матери не было гипертрихоза (и не могло быть, так как у нее нет Y - хромосомы), но она была шестипалой. Следовательно, у нее должен быть хотя бы один ген шестипалости – А. В этой семье родилась нормальная девочка. Ее генотип ХXаа. Один ген пятипалости она получила от отца, а второй ген пятипалости могла получить только от матери. На основе этого решаем, что мать была гетерозиготна по гену шестипалости. Ее генотип ХХАа. Вероятны генотипы детей:

Без обоих аномалий возможна лишь 1/4 детей, или 25%.
Наследование отсутствия потовых желез у человека как рецессивного сцепленного с полом признака
Задача 5
У человека отсутствие потовых желез проявляется как сцепленный с полом рецессивный признак. Глухота, то есть отсутствие слуха, обусловлено аутосомным рецессивным геном. У супружеской пары, нормальной по этим признакам, родился сын с обоими аномалиями. Определите возможные генотип родителей и вероятность рождения ребенка с таким же генотипом как первый. Дать цитологическое обоснование.
Решение:

ак как оба родителя были здоровы, значит ген отсутствия потовых желез является рецессивным и находиться в Х – хромосоме у женщины, т.к. она имеет две Х – хромосомы (в одной Х – хромосоме доминантный ген нормы, а в другой Х – хромосоме – рецессивный ген. Оба родителя гетерозиготы по гену глухоты, потому что у них родился больной сын

Вероятность рождения в этой семье больного ребенка с обоими аномалиями (ааХ b Y) - 1/16 или 0,0625%.
Цитологическое обоснование.
(Гены находятся в хромосомах: - Х-хромосома с генами, - У)

Взаимодействие генов: полное доминирование. Законы генетики, которые использовали при решении задачи: чистоты гамет, сцепленное с полом наследование, закон независимого наследования признаков.
Задача 6
У дрозофилы гены определяющие окраску глаз, локализованы в Х – хромосоме. Доминантный аллель W детерминирует красную окраску глаз, его рецессивный аллель w – белую. Скрещивали гомозиготную красноглазую самку с белоглазым самцом. В F 1 получили 48 потомков. От скрещивания их между собой получено 192 мухи в потомстве F 2 .
Определите:
1. Сколько женских особей было в F 1 ?
2. Сколько самцов в F 1 имело красную окраску глаз?
3. Сколько самок F 1 было красноглазых?
4. Сколько самцов в F 2 было белоглазых?
5. Составить схему скрещивания.
Решение:
Х W – ген определяющий красные глаза
Х w – ген определяющий белые глаза
1). Скрещивание гомозиготной красноглазой самки с белоглазым самцом:

Соотношение особей в потомстве 1:1. По фенотипу все особи красноглазые.
Поэтому:
1. Женских особей в F 1 было 48:2=24особи.
2. В F 1 все самцы (24особи) имели красные глаза.
3. Все самки в F 1 были красноглазыми (24 особи).
2). Скрещивание между собой гибридов F 1:

Поэтому: в F 2 белоглазых самцов было (192:4; х 2 = 96 особей).
Взаимодействие генов: полное доминирование. Законы генетики: закон чистоты гамет и сцепленное с полом наследование.
Задача 7
Селекционеры в некоторых случаях могут определить пол только что вылупившихся цыплят.
При каких генотипах родительских форм, возможно это сделать, если известно, что гены золотистого (коричневого) и серебристого (белого) оперения расположены в Х – хромосоме и ген золотистого оперения рецессивен по отношению к серебристому? Не забудьте, что у кур гетерогенным полом является женский!
Решение:
А – ген серебристого окрашивания
а – ген золотистого окрашивания

При скрещивании серебристой курочки с золотистым петушком курочки будут все золотистые, а петушки серебристые. Таким образом, петушки с серебристой окраской оперения будут иметь генотип Х A Х a , а курочки с золотистой окраской оперения - Х A Y.
Задача 8
Гипертрихоз наследуется как сцепленный с У – хромосомой признак, который проявляется лишь к 17 годам жизни. Одна из форм ретинита (ночная слепота) наследуется как рецессивный, сцепленный с Х – хромосомой признак. В семье, где женщина по обоим признакам здорова, а муж является обладателем только гипертрихоза, родился мальчик с ретинитом. Определить вероятность проявления у этого мальчика гипертрихоза. Определить вероятность рождения в этой семье детей без обоих аномалий и какого, они будут пола.
Решение:
X A – ген нормального ночного зрения;
Х a –ген ночной cлепоты;
Y* - ген гипертрихоза;
Y – ген нормы

Соотношение особей в потомстве 1:1:1:1
Вероятность проявления у первого сына гипертрихоза – 100%. Вероятность рождения здоровых детей – 50% (они будут только девочки).
Задача 9
У человека есть несколько форм стойкого рахита. Одна из его форм наследуется доминантно сцеплено с полом, вторая рецессивно – аутосомная. Какова вероятность рождения больных детей, если мать гетерозиготная по обоим формам рахита, а отец здоровый все его родственники здоровы?
Решение:
Х А – рахит (первая форма);
Х a – норма;
В – норма;
b – рахит (вторая форма).
Из условия задачи видно, что генотип женщины X А X a Bb, а мужчины - X a YBB – он гомозиготен по второй паре генов, т.к. все его родственники здоровы.
Вероятность больных детей 4/8 или 50%. Взаимодействие генов: полное доминирование. Законы генетики, которые использовали при решении задач: закон чистоты гамет, сцепленное с полом наследование, закон независимого комбинирования признаков.
Задача 10
У некоторых пород кур гены, определяющие белый цвет и полосатую окраску оперения, сцеплены с Х – хромосомой, полосатость доминирует над белой сплошной окраской. Гетерогаметный пол у кур женский. На птицеферме белых кур скрестили с полосатыми петухами и получили полосатое оперение как у петушков, так и у кур. Затем скрестили особи, полученных от первого скрещивания, между собой и получили 594 полосатых петушка и 607 полосатых и белых курочек. Определите генотипы родителей и потомков первого и второго поколения.
Решение:
Х А – полосатые;
Х a – белые
У кур гетерогаметный пол, у петухов гомогаметный. Если в F 1 все потомки независимо от пола полосатые, то петух гомозиготен и полосатость – доминантный признак. Во втором поколении наблюдается расщепление признака, поэтому петушок в F 2 будет гетерозиготен.
a)

Половое размножение свойственно всем живым организмам за исключением тех, которые вторично утратили половой процесс. Определение и развитие пола — сложный процесс, который детерминирован генетически, т.е. находится под контролем генов, а также подвержен влиянию внешней среды.

В животном мире господствует раздельнополость, т.е. существуют два типа ясно различающихся в половом отношении организмов — самцы и самки. Различия между ними очень глубокие и затрагивают не только органы, непосредственно участвующие в половом размножении. Половые различия сопровождаются заметными различиями в росте, обмене веществ, инстинктах, а также в тех признаках, которые подвержены воздействию половых желез, например, гребни, рога, волосы, оперение.
Гермафродитизм у животных в норме встречается только у немногих видов, например у червей.
У растений, наоборот, преобладает гермафродитность . Половые различия у растений выражены менее резко, чем у животных. Для растений характерны переходы от обоеполости к однополости, частые аномалии в развитии генеративных органов, изменение пола под влиянием внешних условий.
Определение пола у разных организмов может происходить на разных стадиях жизненного цикла.
Пол зиготы может предопределяться еще в процессе созревания женских гамет — яйцеклеток. Такое определение пола называется прогамным , т.е. оно происходит до оплодотворения. Прогамное определение пола обнаружено у коловраток и кольчатых червей. Яйцеклетки этих животных в результате неравномерного распределения цитоплазмы в процессе оогенеза различаются по размеру. Из крупных яйцеклеток после определения развиваются только самки, из мелких — только самцы.
Наиболее распространенным типом определения пола является сингамное , т.е. определение пола в момент слияния женских и мужских гамет. Оно встречается у млекопитающих, птиц, рыб и др.
Известен также третий тип определения пола — эпигамное , которое происходит на ранних стадиях индивидуального развития особи (например, у морского червя Bonelia viridis).
У большинства животных и раздельнополых растений основную роль в определении пола играют половые хромосомы . Еще в начале ХХ в. (1902 г., McClung) было установлено, что у некоторых насекомых (клоп Protenor) самцы образуют два типа сперматозоидов: один тип — с лишней хромосомой, второй — без нее. У самцов клопа Protenor в одних сперматозоидах было 7 хромосом, в других — 6. Непарную хромосому назвали половой хромосомой, в отличие от остальных — аутосом . В соматических клетках самца содержится 13 хромосом, одна из которых Х-хромосома (12A+X), в соматических клетках самки — 14 хромосом (12A+XX). Женский пол клопа является гомогаметным, так как образует гаметы одного типа (6A+X), а мужской — гетерогаметным и образует два типа гамет (6A+X) и (6А+0). Такой тип определения пола, при котором самки имеют кариотип ХХ , а самцы — Х0 , назван Protenor-типом. Он описан у большинства прямокрылых насекомых, жуков, пауков, многоножек и нематод.
Вслед за Protenor-типом был открыт другой тип определения пола, который характерен для млекопитающих, многих рыб, амфибий и ряда растений. Впервые он был описан у клопа Lygaeus turcicus и получил название Lygaeus-типа. При этом типе определения пола имеются два вида половых хромосом: Х и Y . Самки имеют две хромосомы, а самцы одну Х-хромосому и непарную ей Y-хромосому. Обозначение половых хромосом буквами X и Y отражает их форму, которую они имеют в профазе мейоза в результате отталкивания хроматид, соединенных только в области первичной перетяжки.
Женский пол при типе Lygaeus является гомогаметным, мужской — гетерогаметным.
У птиц, некоторых видов бабочек и рыб тип определения пола — обратный Lygaeus, т.е. гомогаметным является мужской пол. В этом случае для обозначения половых хромосом используют другие буквы: ♀ZW, ♂ZZ.
У моли описан тип — обратный Protenor, т.е. ♀Х0, ♂ХХ.
Особый тип определения пола характерен для пчел. Здесь разница между полами затрагивает не одну пару хромосом, а весь набор. Самки пчел — диплоидны, а самцы — гаплоидны, так как женские особи развиваются из оплодотворенных яйцеклеток, мужские особи — в результате партеногенеза.
Хромосомный механизм определения пола у растений был впервые определен у печеночного мха — Sphaerocarpus в ходе тетрадного анализа. Из четырех спор, образующихся в результате мейотического деления материнской клетки, две дают начало женским растениям, а две другие — мужским. Поскольку хромосомы мха Х и Y морфологически легко различимы, было установлено, что женские растения имеют кариотип 7А + Х, а мужские — 7А + Y. Диплоидный спорофит, который образуется в результате оплодотворения, имеет кариотип 14А + XY.
Гетероморфные пары хромосом обнаружены у мужских растений дремы, конопли, щавеля, хмеля и др. Определение пола у них соответствует типу Lygaeus. У земляники гетерогаметным (XY ) является женский пол, мужской — гомогаметным.

Половые хромосомы отличаются от аутосом поведением в профазе мейоза. Во время гаметогенеза они находятся в сильно спирализованном состоянии и редко объединяются в биваленты. Тем не менее они обладают сегментной гомологией и проявляют тенденцию к частичной коньюгации.
X и Y -хромосомы различаются по форме, величине и генному составу. Х-хромосома чаще всего относится к разряду крупных хромосом с большим генетическим объемом. У дрозофилы Х-хромосома — самая крупная в наборе. У человека Х-хромосома относится к разряду средних метацентриков, с нарушением ее структуры связан ряд тяжелых наследственных патологий (синдромов). Мужскую половую хромосому характеризует обедненность генами и, соответственно, низкая генетическая активность, а иногда и полная инертность. У человека с помощью молекулярно-генетических методов в Y-хромосоме выявлено около 40 генов. Однако реальных генетических функций еще меньше. В частности, в Y-хромосоме лежит мутация, отвечающая за малоприятный для мужчин признак — волосатость ушей. У дрозофилы Y-хромосома практически не оказывает никакого влияния на развитие пола.
У растений Y-хромосома также ведет себя по-разному: у одних она играет активную роль в определении пола, у других — является инертной. Например, Y-хромосома Milandrium alba (дрема) имеет сегменты, потеря которых ведет к нарушению нормального процесса развития пола и, как следствие, к мужской или женской стерильности. У Rumex acetosa Y-хромосома генетически инертна. У некоторых растений активность Y-хромосомы настолько высока, что особи YY оказываются жизнеспособными, как у аспарагуса, в то время как у других видов подобные особи не выживают.
Если гены, детерминирующие признаки, находятся в половых хромосомах, то их наследование не подчиняется законам Менделя. Распределение этих признаков соответствует распределению половых хромосом в процессе мейоза. Поскольку большинство генов, локализованных в Х-хромосоме, не имеют своих аллелей в Y-хромосоме, то у гетерогаметного пола (XY) в фенотипе проявляются все рецессивные гены, содержащиеся в их единственной Х-хромосоме. Гены, если они имеются в Y-хромосоме, проявляются также только у гетерогаметного пола.
Наследование признаков, определяемых генами, локализованными в Х и Y-хромосомах, называют сцепленным с полом. Впервые оно было описано Т. Морганом и его коллегами на примере рецессивного признака “white” — белые глаза.
Как видно из схемы, результаты прямого и обратного скрещиваний в случае наличия сцепления с полом разные. В прямом скрещивании гомозиготная красноглазая самка передает доминантный ген W и дочерям и сыновьям, благодаря чему все гибриды F 1 имеют красные глаза. Скрещивание гетерозиготных самок F 1 с самцами F 1 дает в F 2 только красноглазых самок, одна половина которых является гомозиготными, а другая — гетерозиготными. Среди самцов F 2 наблюдается расщепление на красноглазых и белоглазых в соотношении 1: 1, которое обусловлено гетерозиготностью самок F 1 , так как свою единственную Х-хромосому сыновья наследуют от матери. Общая формула расщепления по окраске глаз в F 2 (без учета пола) — 3: 1. На наличие сцепления признака с полом указывает то, что белая окраска глаз в F 2 проявляется только у самцов.

В обратном скрещивании рецессивная гомозиготная белоглазая самка передает ген w вместе с Х-хромосомой и дочерям и сыновьям F 1 , но проявляется он только у самцов. У самок F 1 этот ген подавляется доминантным аллельным геном, полученным от отца, и поэтому глаза у них красные. Таким образом, признак передается от отца к дочерям, а от матери к сыновьям. Такое наследование называется крисс-кросс (крест-накрест). Скрещивание самок и самцов F 1 дает мух двух фенотипических классов (красноглазых и белоглазых) в соотношении 1: 1, которое полностью соответствует распределению половых хромосом.
Описанный тип наследования окраски глаз у дрозофилы является закономерным для всех организмов в отношении признаков, которые определяются генами, локализованными в Х-хромосоме.

Сцепленное с полом наследование используется для ранней диагностики пола у животных, что важно для сельскохозяйственного производства. В птицеводстве важно определять пол “суточных” цыплят, чтобы ставить петушков и курочек на разный рацион, откармливая петушков на мясо. Для диагностики пола используется крисс-кросс наследование признака окраски пера. При скрещивании пестрой курицы (признак доминантный) с черным петухом (признак рецессивный) в F 1 все петушки, получившие доминантный ген от матери, будут пестрыми, а курочки — черными.
У человека сцепленно с полом наследуются такие наследственные аномалии, как гемофилия и дальтонизм. Поскольку у человека гетерогаметным является мужской пол, то эти аномалии проявляются, в основном, у мужчин. Женщины обычно являются носительницами таких генов, имея их в гетерозиготном состоянии.
При разведении тутового шелкопряда крисс-кросс наследование используется для отбора самцов по окраске грены (признак сцеплен с полом), так как выход шелка из коконов тутового шелкопряда мужского пола на 20-30% выше.
Картина сцепленного с полом наследования может искажаться, если наблюдаются отдельные случаи нерасхождения половых хромосом в процессе мейоза. Так, при скрещивании белоглазой самки дрозофилы с красноглазым самцом (см. выше схему наследования крисс-кросс) в F 1 , помимо красноглазых самок и белоглазых самцов, появляются единичные белоглазые самки и красноглазые самцы. Причиной этого отклонения является нерасхождение Х-хромосом у исходной самки. В процессе гаметогенеза в яйцеклетку попадает не одна Х-хромосома, а обе, или же, наоборот, ни одной, а обе попадают в полярное тельце. При оплодотворении таких яйцеклеток нормальными сперматозоидами и развиваются красноглазые самцы и белоглазые самки.
Потомство, которое образуется в результате первичного нерасхождения хромосом у самки, имеет разные, не соответствующие норме сочетания и количество половых хромосом. Однако, генетическая инертность Y-хромосомы делает особей с кариотипом ХХY женскими и жизнеспособными, а с кариотипом Х0 — мужскими и также жизнеспособными. Зиготы, не получившие Х-хромосомы (Y0 ), погибают, так же как (за редким исключением) и зиготы с тремя Х-хромосомами.
Схема наследования белой окраски глаз у дрозофилы (ген white)
при нерасхождении X-хромосом у самки

У дрозофилы выведена линия (double yellow — двойная желтая), у которой из поколения в поколение нарушается наследование сцепленного с полом признака — желтая окраска тела. У самок этой линии Х-хромосомы соединены друг с другом в проксимальной части и имеют одну центромеру. В связи с этим в мейозе они ведут себя как одна хромосома и в анафазе отходят к одному полюсу.
Гетерогаметность одного пола определяет соответствие соотношения полов в каждом поколении организмов формуле 1: 1. Это соотношение совпадает с расщеплением при анализирующем скрещивании. Рассмотрим его на примере дрозофилы, у которой определение пола соответствует Lygaeus-типу. Набор хромосом у дрозофилы состоит из трех пар аутосом и двух половых хромосом. Самка образует один тип гамет с гаплоидным набором (3A+X), а самец в равных количествах два типа гамет (3A+X) и (3A+Y). В итоге в следующем поколении развивается одинаковое количество самок и самцов.

Такое наследование наблюдается при разных типах хромосомного механизма определения пола, и вероятность рождения потомков мужского и женского пола в норме одинакова. Однако баланс полов может быть нарушен, если в половых хромосомах возникают летальные мутации. Рассмотрим случай, когда рецессивная летальная мутация (l ) возникла в одной из двух Х-хромосом самки дрозофилы (X Bl ), маркированной доминантной мутацией Bar (В ) — полосковидные глаза. Рассмотрите схему скрещивания такой самки с нормальным самцом дикого типа (+), имеющим круглые глаза.

Как видно из схемы, появление рецессивной летальной мутации в одной из Х-хромосом самки приводит к гибели половины мужского потомства. Об этом судят по отсутствию самцов с полосковидными глазами, получившими от матери Х-хромосому с летальным геном (X Bl ).
Гены, определяющие признаки пола, имеются не только в половых хромосомах, но и в аутосомах. С другой стороны, признаки, которые наследуются сцепленно с полом, часто не имеют прямого отношения к полу. Существует особая категория признаков, которые проявляются только у одного пола. Это — ограниченные полом признаки . Определяющие их гены имеются у обоих полов и могут находиться как в половых хромосомах, так и аутосомах. Однако работают эти гены, т.е. проявляют свое действие на уровне фенотипа, только у одного пола. К числу таких признаков относятся, например, молочность и жирность молока у коров, яйценоскость и размер яиц у кур. Эти признаки, которыми обладают особи женского пола, могут целиком определяться генотипом отца. Такое явление широко используется в селекции животных при использовании отцовских особей-производителей для получения высококачественного потомства.
Гены, определяющие развитие вторичных половых признаков, имеются как у мужчин, так и у женщин, но их проявление контролируется гормонами.
Пол может оказывать влияние на характер проявления признака, т.е. на его доминантность или рецессивность. В этом случае признаки называют зависимыми от пола . Так, например, у овец ген, определяющий развитие рогов, является доминантным у самцов и рецессивным — у самок. В связи с этим гетерозиготные самки являются комолыми, а гетерозиготные самцы — рогатыми. У человека точно так же наследуется признак плешивости. Зависимые от пола признаки находятся под сильным влиянием половых гормонов, соотношение которых может либо усилить, либо ослабить экспрессию гена.
Итак, подведем итог, касающийся механизма определения пола. Пол, как любой другой признак организма, детерминирован генетически. В определении пола у большинства животных и растений основная роль принадлежит половым хромосомам. Расщепление по полу соответствует соотношению 1: 1, что обусловлено равновероятным образованием двух типов гамет (1/2 с Х и 1/2 с Y хр.) у гетерогаметного пола (XY ). Гетерогаметным может быть как мужской, так и женский пол.
Определение пола — это начальный этап становления пола, за которым следует процесс его дифференциации, приводящий к развитию двух разных половых типов — женского и мужского. У животных половая дифференциация затрагивает всю организацию особи: строение органов размножения, внешнюю морфологию, обмен веществ, поведение, гормональный баланс, продолжительность жизни и пр. Половые различия которые обеспечивают комбинативную изменчивость внутри вида, а также его изоляцию, являются адаптивным механизмом.
Различают первичные и вторичные половые признаки. Первые непосредственно обеспечивают осуществление полового процесса. В частности, к ним относятся различия в строении внешних и внутренних половых органов женских и мужских особей. Развитие вторичных половых признаков является результатом нормального функционирования гонад (т.е. опосредовано первичными половыми признаками) и способствует половому размножению. Регулируется развитие вторичных половых признаков с помощью половых гормонов.
На процесс дифференциации пола оказывают влияние как генотипические факторы, так и внешняя среда.
Еще в начале ХХ в. было высказано предположение, что зигота является потенциально бисексуальной, но существуют механизмы, осуществляющие дифференциацию пола. Одним из таких механизмов является баланс половых хромосом и аутосом, при нарушении которого развитие пола отклоняется либо в сторону женского, либо в сторону мужского пола. Необходимость такого баланса впервые была установлена в опытах К. Бриджеса (лаборатория Т. Моргана), который обнаружил линию дрозофилы, дающую наряду с нормальными самцами и самками большой процент интерсексов. Интерсексы представляют собой смесь первичных и вторичных мужских и женских половых признаков, образуя все переходные типы: от сходных в основном с самцами до сходных с самками. Все они стерильны. В опыте Бриджеса они возникли в потомстве триплоидных самок, оплодотворенных нормальными диплоидными самцами, и содержали три набора аутосом и нормальное количество половых хромосом: 2Х+3А. Наряду с типичными интерсексами, в потомстве были представлены особи с гипертрофированными признаками женского пола — суперсамки (3Х+2А), и мужского пола — суперсамцы (XY+3X).
На основании этих результатов Бриджес пришел к выводу, что не само присутствие двух половых хромосом (XX или XY) определяет развитие пола, а баланс половых хромосом и гаплоидных наборов аутосом. Поскольку у дрозофилы Y-хромосома генетически инертна, то важно только количество Х-хромосом. Все особи с отношением 2Х: 2А = 1 являются самками, особи с отношением 1Х: 2А = 0,5 — самцами, типы с промежуточными между 1 и 0,5 отношениями являются интерсексами, а отношения больше 1 дают суперсамок, меньше 0,5 — суперсамцов.
Аномальное развитие пола при изменении числа наборов аутосом обусловлено нарушением баланса генов, которые участвуют в развитии пола. Поскольку гены проявляют свое действие в конкретных условиях, то на их функционирование оказывают влияние внешние факторы. Так, потомство триплоидных самок дрозофилы воспитывалось в условиях высокой и низкой температур. В обоих случаях развивались интерсексы, но при высокой температуре преимущественно с признаками самки, а при пониженной — с признаками самца. Таким образом, окончательное развитие пола является результатом сложных взаимодействий генов, локализованных как в половых хромосомах, так и в аутосомах, друг с другом и с факторами окружающей среды.
Изначальная бисексуальность зигот подтверждается фактами переопределения пола в процессе индивидуального развития. Классический пример — морской червь Bonellia viridis. Свободноплавающие личинки этого червя развиваются в самок. Если же личинка остается прикрепленной к материнской особи, из нее развивается самец. Будучи отделена от самки, такая личинка, начавшая развиваться в самца, изменяет направление дифференциации пола в женскую сторону и из нее развивается интерсекс. В хоботке самки имеются химические регуляторы, способные переопределять пол личинок.

Большой интерес представляет экспериментальное переопределение пола. Путем воздействия гормональными препаратами у ряда животных удается получить полное превращение пола вплоть до способности формировать половые клетки противоположного пола. Такое превращение известно у некоторых лягушек, рыб, птиц и других животных. Так, раннее удаление яичника у самок кур и голубей может изменить в мужскую сторону окраску оперения, поведение и даже вызвать развитие семенника. У крупного рогатого скота наблюдались случаи рождения разнополых двойнь, в которых бычок оказывался нормальным, а телка — стерильной, со многими чертами самцового типа. Такие двойни носят название “фримартинов”. Их появление обусловлено тем, что семенники мужского эмбриона рано начинают выделять мужской гормон, который попадает в кровь и оказывает влияние на близнеца.
Один из ярких примеров полного переопределения пола описан в 1953 г. японским ученым Т. Ямамото. Опыт проводился на белых и красных медаках (Oryzias latipes), у которых доминантный ген красной окраски находится в Y-хромосоме. При такой локализации гена при скрещивании самцы всегда будут красными, а самки — белыми. Фенотипических самцов кормили с добавлением в корм женского полового гормона. В результате оказалось, что все красные рыбки с генотипом самца являются самками с нормальными яичниками и женскими вторичными половыми признаками.
Переопределение пола может быть следствием мутаций отдельных генов, участвующих в дифференциации пола. Так, у дрозофилы в одной из аутосом обнаружен рецессивный ген tra , присутствие которого в гомозиготном состоянии обусловливает развитие женских зигот (XX) в фенотипических самцов, оказывающихся стерильными. Самцы XY, гомозиготные по этому гену, являются плодовитыми.
Аналогичные гены найдены у растений. Так, у кукурузы рецессивная мутация silkless в гомозиготном состоянии вызывает стерильность семяпочек, в связи с чем обоеполое растение функционирует как мужское. У сорго обнаружены два доминантных гена, комплементарное взаимодействие которых также вызывает женскую стерильность.
У наездника Habrobracon пол определяется по тому же типу, что и у пчел: диплоидные самки развиваются из оплодотворенных яиц, а гаплоидные самцы партеногенетически. Но иногда самцы могут развиваться из оплодотворенных яиц. Причина такой ситуации лежит в действии специфического гена, в гомозиготном состоянии определяющего развитие зиготы по мужскому типу.
Правильность хромосомной теории определения пола подтверждается существованием половых мозаиков, или гинандроморфов , совмещающих в себе части тела мужского и женского полов. Известны разные типы гинандроморфов: латеральные, переднезадние, мозаичные.

Билатеральный гинандроморф
Drosophila melanogaster
Латеральный гинандроморфизм описан у насекомых, у кур, у певчих птиц. В этом случае одна половина тела соответствует женскому типу, вторая — мужскому. При мозаичном гинандроморфизме большая часть тела имеет признаки одного пола, и лишь отдельные участки — признаки противоположного пола. Этот тип описан, в частности, у дрозофилы. Чаще всего причиной появления гинандроморфов является утрата одной из двух Х-хромосом в раннем дроблении зиготы с кариотипом самки (ХХ). Клетки с кариотипом Х0 обнаруживают признаки мужского пола. Чем раньше произойдет элиминация Х-хромосомы, тем больше участков мужского типа будет представлено в теле взрослой мухи. Обнаруживаются такие мозаики по проявлению рецессивных генов, которые в зиготе находились в гетерозиготном состоянии, но проявились фенотипически в клетках с кариотипом Х0.
Еще одной причиной гинандроморфизма может быть развитие зародыша из яйцеклетки с двумя ядрами (дизиготический гинандроморфизм). В этом случае мозаики могут быть соматическими, если оба ядра имеют один и тот же набор половых хромосом, но разный генотип (например, одно ядро Аа, а другое — аа), или половыми, если одно ядро ХХ, а другое ХY, или теми и другими одновременно. Подобный тип гинандроморфизма описан у шелковичного червя, бабочки, дрозофилы.
Известен также гинандроморфизм, причиной которого является полиспермия. Он обнаружен у дрозофилы. В яйцеклетке дрозофилы могут сформироваться два женских гаплоидных пронуклеуса, с одной Х-хромосомой каждый. При проникновении в яйцеклетку двух сперматозоидов один пронуклеус может оплодотвориться сперматозоидом с Х-хромосомой, а другой — сперматозоидом с Y-хромосомой. После первого дробления образуются два бластомера, один с кариотипом ХХ, другой — ХY, что в дальнейшем приведет к развитию гинандроморфа.
Передача какого-либо гена, находящегося в половых хромосомах, от родителей потомству называется наследованием, сцепленным с полом. Переданные признаки проявляются у потомков определённого пола.
Половые хромосомы
Определение сцепленного с полом наследования взаимосвязано с понятием половых хромосом.
Все хромосомы делятся на два вида:
- аутосомы - одинаковые для обоих полов;
- гоносомы - половые хромосомы, определяющие пол.
Организм развивается в соответствии с полом (мужским или женским), который несёт половая хромосома (обычно пара). Различать организмы по половому признаку можно только в случае полового размножения - оогамии.
С точки зрения генетики выделяют два типа пола:
- гомогаметный - содержит одинаковые хромосомы;
- гетерогаметный - содержит разные хромосомы.

Рис. 1. Гомогаметный и гетерогаметный пол.
У людей и других млекопитающих гомогаметный пол - женский, гетерогаметный - мужской. Женские хромосомы условно обозначают XX, мужские - XY. У птиц наоборот: женский пол - гетерогаметный (обозначается ZW), а мужской - гомогаметный (ZZ). Некоторые животные имеют несколько пар хромосом, определяющих пол.
Генотип человека по половому признаку выглядит следующим образом:
- мужчина - 22 пары аутосом и XY;
- женщина - 22 пары аутосом и XX.

Рис. 2. 23 пара хромосом.
ТОП-2 статьи которые читают вместе с этой
Механизм наследования
При оплодотворении пол определяет Х-хромосома. Если данная хромосома достаётся от отца, то рождается девочка, если от матери - мальчик. Y-хромосома несёт только пол и не сцеплена с каким-либо признаком. С Х-хромосомой наследуется около 60 генов, отвечающих не только за признаки, но и различные заболевания.
В частности Х-хромосома может нести:
- гемофилию - заболевание, связанное с нарушением свёртываемости (коагуляции) крови;
- дальтонизм - нарушение восприятия цветов (болеющий человек часто путает красный и зелёный цвета);
- потемнение эмали зубов;
- мышечную дистрофию.
Рассмотрим наследование признаков, сцепленных с полом , на примере гемофилии:
- в Х-хромосоме может находиться доминантный ген (H), отвечающий за нормальную свёртываемость крови, или рецессивный (h), отвечающий за гемофилию;
- если женщина гетерозиготна (XHXh), но гемофилия у неё не проявляется, но она является носителем болезни, т.к. содержит рецессивный ген;
- мужчина, несущий только одну Х-хромосому, может содержать только один из вариантов генов - H или h, поэтому даже при наличии рецессивного гена мужчина заболевает гемофилией (ген свёртываемости отсутствует).

Рис. 3. Наследование гемофилии.
Если мать гетерозиготна, а отец болеет гемофилией, то шансы родиться больными у девочек и мальчиков уравниваются (вероятность 50 %).
Передача наследственного признака от гетерозиготной матери и больного отца выглядит следующим образом:
XHXh х XhY
F1: XHXh (здоровая дочь) XhXh (больная дочь)
XHY (здоровый сын) XhY (больной сын)
Если женщина гомозиготна, то дети родятся здоровыми, даже если отец болен гемофилией:
XHXH х XhY
F1 2XHXh 2XHY
У кошек черепаховый (трёхцветный) окрас сцеплен с Х-хромосомой. Необычный окрас проявляется, когда кошка имеет гены, одновременно отвечающие за рыжую и чёрную окраску (XAXB). Кот же может получить только один из генов - рыжий (XAY) или чёрный (XBY).
Что мы узнали?
Узнали о том, что понимают под наследованием, сцепленным с полом. Признаки несут и передают обоим полам только Х-хромосомы. У женщин меньше шансов получить врождённую болезнь, сцепленную с полом, чем у мужчин, т.к. женщины содержат две Х-хромосомы, а мужчины - только одну.
Тест по теме
Оценка доклада
Средняя оценка: 4.4 . Всего получено оценок: 153.
Решения задач
Хромосомная теория наследственности объяснила загадку рождения организмов разного пола. Она установила, что пол наследуется, как менделирующий признак, сочетание половых хромосом в зиготе определяет пол будущего организма. У всех млекопитающих (в том числе и у человека), у дрозофилы и многих других животных женские особи в соматических клетках имеют две X-половые хромосомы, а мужские Х- и У- хромосомы. Женский пол является гомогаметным и образует яйцеклетки одного типа с Х- половой хромосомой. Мужской пол является гетерогаметным и образует в равном количестве сперматозоиды двух типов: одни содержат Х-половую хромосому, другие - Y-половую хромосому. При оплодотворении возможны две комбинации. Ниже приводится схема наследования пола у человека. А - гаплоидный набор аутосом, 2А - диплоидный набор аутосом, X и Y - половые хромосомы.
Если в зиготе встречаются две Х-половые хромосомы, то развивается женская особь; если в зиготе сочетаются Х- и Y-хромосомы, то из такой зиготы развивается мужская особь. Число особей женского пола (2А + XX) относится к числу особей мужского пола (2А + ХУ) как 1:1. Фактически же на 100 девочек в России рождается 102-103 мальчика, в США - 106, Греции - 113. Это сингамное определение пола, при котором пол будущей особи детерминируется в момент слияния гамет. Известны еще два способа определения пола: эпигамный (червь бонеллия) и прогамный. При прогамном определении пола - пол будущего организма определяется еще во время формирования женских гамет. Из крупных яйцеклеток после оплодотворения развиваются самки, из мелких - самцы (тли, некоторые кольчатые черви).
В противоположность млекопитающим, у птиц и некоторых видов бабочек женский пол является гетерогаметным, а мужской - гомогаметным. Сперматозоиды петуха все одинаковые и содержат Х-половую хромосому, яйцеклетки бывают двух типов: одни содержат Х-половую хромосому, другие - Y-половую хромосому. У некоторых насекомых (прямокрылые, ручейники) Y-половая хромосома отсутствует, в таком случае кариотип самцов 2А + ХО, самок 2А +
XX. Таким образом, в природе встречаются следующие типы половых хромосом:
1. ♀XX, ♂XY класс млекопитающих, класс насекомых - мушка дрозофила и др.
2 ♀XY, ♂XX (♀ZW, ♂ZZ) класс насекомых, (бабочки), класс птицы, класс пресмыкающиеся, класс двудольных растений, семейство розоцветных, некоторые сорта земляники
3. ♀XX, ♂ХО класс насекомых, отряды прямокрылых, ручейников.
Таким образом, у всех организмов пол определяется наследственными факторами и только в некоторых случаях на процесс формирования мужских или женских особей решающее влияние оказывают факторы внешней среды. Наследование признаков, сцепленных с полом.
Признаки, наследуемые через половые хромосомы, получили название сцепленных с полом. У человека сцепленно с Х-половой хромосомой по рецессивному типу наследуются гемофилия (наследственное заболевание, характеризующееся повышенной кровоточивостью вследствие недостатка факторов свертываемости крови), дальтонизм, мышечная дистрофия Дюшена и другие; по доминантному типу - темная эмаль зубов, витаминоустойчивый рахит и другие (267 генов локализовано в Х-хромосоме, данные на 1985 г., Мак-Кьюсик). Признаки, наследуемые через Х-хромосому, встречаются как у лиц мужского, так и женского пола. Женщина может быть как гомо-, так и гетерозиготна по генам, локализованным в Х-хромосоме. Рецессивные же аллели гена в Х-половой хромосоме у женщины проявляются только в гомозиготном состоянии. Поскольку у лиц мужского пола только одна Х-хромосома, все локализованные в ней гены, даже рецессивные, сразу же проявляются в фенотипе. Такой организм называется гемизиготным, так как в Y-половой хромосоме не имеется гомологичных локусов Х-половой хромосоме. За редким исключением аллелей генов общей цветовой слепоты, пигментного ретинита и других. Особенности наследования с Х-половой хромосомой
Если признак рецессивен, то мужчины его наследуют чаще, чем женщины. Обычно женщина бывает проводником (кондуктором) данного признака, девочки наследуют такой признак только от отца. Если в браке оба родителя фенотипически здоровы, но женщина - носительница рецессивного признака, этот признак наследуется у 50% сыновей, и половина дочерей являются носителями рецессивного признака.
Признаки, которые наследуются через Y-половую хромосому (гипертрихоз - волосатость мочки уха и ушной раковины, сращение кожных покровов I и III пальцев стопы и другие) получили название голандрических, они могут быть только у лиц мужского пола и передаются от отца к сыну, от сына к внуку, правнуку и так далее.
Примеры обозначения:
X - нормальная свертываемость крови,
X h - гемофилия,
У 1 -гипертрихоз,
X D - нормальное цветовое зрение,
X d - дальтонизм,
Частично сцепленное с полом наследование:
Х А - нормальное цветовое зрение,
У А - нормальное цветовое зрение,
Х а - общая цветовая слепота,
У а - общая цветовая слепота,
X H X h - генотип женщины-носительницы гемофилии,
X h Y - генотип мужчины - гемофилика,
XY r - генотип мужчины, имеющего гипертрихоз,
X d X d - генотип женщины - дальтоника,
X d Y - генотип мужчины - дальтоника,
X A Y a ; X a Y a ; X a Y A - генотипы с общей цветовой слепотой.
Задача 1.
На определение генотипа и фенотипа потомства при сцепленном с полом наследовании.
Девушка, имеющая нормальное зрение, отец которой страдал цветовой слепотой (дальтонизм), выходит замуж за здорового мужчину, отец которого также был дальтоником. Дальтонизм наследуется рецессивно сцепленно с X половой хромосомой. Какое зрение можно ожидать у детей в данном браке?
Решение
X D - нормальное цветовое зрение
X d - дальтонизм
Определите F по зрению?
Женщина в браке, имела отца-дальтоника, следовательно по генотипу она гетерозиготна X D X d и носительница аномального гена дальтонизма, так, как одну Х- половую хромосому в зиготе дочь получает от отца, а другую Х- половую хромосому - от матери. Поскольку девушка по условию задачи имела нормальное зрение, следовательно от матери она унаследовала нормальный ген. Мужчина в браке имеет также нормальное зрение, отец которого страдал дальтонизмом. Сын от отца унаследовал У- половую хромосому, в которой нет локуса на ген дальтонизма, мать жениха была здоровой, генотип мужчины в браке Х D У,
Записываем схему брака:

Здоровых - 75%, 25% - страдают дальтонизмом. Исследуя потомство, устанавливаем 75% детей будут здоровыми, все дочери по фенотипу имеют нормальное зрение, так как у отца нет аномального гена, но половина дочерей - носительницы дальтонизма, так как аномальный ген они унаследуют от матери; 25% сыновей всего потомства, или 50% от мужского пола с нормальным цветовым зрением; 25% сыновей от всего потомства и 50% от лиц мужского пола страдают дальтонизмом. По полу дальтониками могут быть только мужчины, так как они гемизиготны по генотипу, получая аномальный рецессивный ген от матери, который несомненно у них проявится в фенотипе. По условию задачи мать фенотипически здорова, но носительница, поэтому вероятность мужчин-дальтоников составляет 50% от лиц мужского пола.
Ответ:_в данном браке вероятность рождения здоровых детей 75%, среди них все девочки, фенотипически здоровые (50%), но половина из них - носительниц гена дальтонизма, здоровых мальчиков 25% и 25% мальчиков от всего потомства и 50% от лиц мужского пола будут дальтониками.
Казанцева Л.В., учитель биологии и химии
Задачи урока.
Сформировать знания о хромосомном механизме определения пола, аутосомах и половых хромосомах, о сцепленном с полом наследовании признаков.
Объяснить возникновение некоторых наследственных болезней человека, связанных с полом.
Продолжить формирование навыков решения генетических задач.
Презентация
Таблица «Хромосомный механизм определения пола»
I этап. Повторение и проверка знаний основных понятий генетики, закономерностей.
Совсем недавно мы начали знакомиться с удивительной наукой-генетикой .
Генетика – это молодая наука, составляющая основу современной биологии, прочно вошедшая в человеческое общество, разгадавшая геном человека, поднявшая на новый уровень современную медицину. Сегодня нет ни одной области деятельности человека, где необходимы знания этой науки. Давайте вспомним ранее изученный материал.
Терминологический диктант
(слайд 1)
Генетика - наука о закономерностях наследственности и изменчивости.
Генотип-совокупность генов организма.
Фенотип-совокупность внешних и внутренних признаков организма.
Аллельные гены- гены, ответственные за развитие одного признака.
Гетерозиготы - организмы, содержащие разные аллельные гены.
Доминантный признак - преобладающий.
Дигибридное скрещивание- скрещивание родительских особей, отличающихся по двум признакам.
Перекрест - обмен аллельными генами между двумя гомологичными хромосомами.
Локус - строго определенное место гена.
Закон сцепления - гены, находящиеся в одной хромосоме наследуются ,как правило,вместе,т.е. сцеплено.
Мы изучили многие генетические закономерности . Сегодня на уроке мы должны ответить на вопросы (слайд 2) : Отчего зависит рождение мужских и женских особей? Почему некоторые болезни женщины передают своим сыновьям, а дочери этими болезнями не страдают? Почему у животных определенные признаки также зависят от пола? Почему соотношение по полу (рождение девочек и мальчиков) приближено 1:1?
Записываем тему урока « Генетика пола. Сцепленное с полом наследование”(слайд3)
Определение пола.
В свое время Менделем было отмечено, что внутри вида количество особей мужского пола приблизительно равно количеству особей женского пола.
Такое расщепление у потомства наблюдается в тех случаях, когда скрещиваются гетерозиготная (Аа) и гомозиготная (аа) родительские особи. Г. Мендель предположил, что один из полов гетерозиготен , второй - гомозиготен по гену, который определяет пол организма. Это предположение было подтверждено в начале 20 века, когда Т. Моргану и его соратникам удалось установить, что самки и самцы различаются по набору хромосом.(слайд 4-6)
У каждого вида организмов имеется определенный набор хромосом. (вводятся понятия с использованием таблицы «Хромосомный механизм определения пола» (слайды 7-10)
Кариотип - набор хромосом (общее число, размер, форма), содержащийся в клетках того или иного вида организмов. Хромосомы, одинаковые у обоих полов, называют аутосомами.
Хромосомы, по которым мужской и женский пол отличаются друг от друга, называют половыми хромосомами.
Пол с одинаковыми половыми хромосомами - гомогаметный, а с разными - гетерогаметный.
IV этап. Домашнее задание: параграф 3.10, задача В каком случае у мужчины дальтоника может быть внук дальтоник (слайд 23)
V этап. Итог урока. Выводы (слайд 24) Выводы:
Хромосомы у организмов делятся на аутосомы и половые хромосомы.
Наследование признаков организма, а также некоторых заболеваний у человека, происходит через половые хромосомы, т.е. сцеплено с полом.
У многих видов расщепление по признаку пола происходит в соотношении 1:1
Хромосомная теория наследственности, дала точное и единое объяснение всех явлений наследования при половом размножении.